Basic Sciences

PREGUNTAS
Antes de continuar, trate de responder las siguientes preguntas. Las respuestas se pueden encontrar al final del artículo con la correspondiente explicación. Por favor conteste verdadero o falso.
- El sistema inmune mediado por células:
a. Puede ser responsable de enfermedades autoinmunes
b. Implica la activación de linfocitos en la periferia
c. Induce la proliferación de células plasmáticas
d. Resulta en un incremento en la síntesis de IgA
e. Causa liberación de grandes cantidades de histamina - Con respecto a las células B:
a. Son llamadas así a medida que van madurando en la médula ósea
b. Muestran receptores de similar morfología a la inmunoglobulina producida por dichas células
c. Dan lugar a las células NK
d. Son solo activadas por células T
e. Produce más de un tipo de anticuerpo - Las immunoglobulinas:
a. Son secretadas por las células T
b. La IgA está presente en la membrana de las mucosas
c. Consiste en una cadena pesada y dos cadenas livianas
d. La especificidad para los antígenos depende de las cadenas livianas
e. La IgG es responsable de la degranulación de los mastocitos
Puntos clave.
- La respuesta inmune innata es no específica y destruye a los patógenos mayormente por fagocitosis y activación del complemento.
- El sistema inmune adquirido genera una respuesta específica a determinados patógenos y esta se realiza principalmente a través de células T, células B, inmunoglobulinas y complemento.
- Las células T pueden ser CD8+ (células T citotóxicas) o CD4+ (células T ayudadoras),
- Las células B secretan immunoglobulinas. Estas se fijan al antígeno de los patógenos y los presenta para su destrucción por el complemento o por actividad fagocitaria.
Introducción
La inmunidad describe la capacidad de luchar contra las infecciones y requiere de un sistema inmune altamente funcional. Como anestesiólogos nosotros ponemos a prueba a diario al sistema inmune con la administración de medicamentos, la intubación traqueal o la colocación de líneas intravasculares. Es importante entender los principios básicos de la inmunología como ciencia para reducir el riesgo de nuestros pacientes y entender porque algunos de ellos son más susceptibles a sufrir infecciones.
La finalidad de este primer tutorial es proveer una visión general de la respuesta inmune mientras que el segundo se concentrará en la inmunología en la práctica clínica.
La respuesta inmune puede ser categorizada en 2 sistemas: inmunidad innata e inmunidad adquirida. El sistema de inmunidad innata representa la primera línea de defensa del organismo y comprende barreras físicas y algunas defensas celulares. Los patógenos que evaden el sistema innato se enfrentan subsecuentemente con la respuesta inmune adquirida, la cual es un sistema específico de respuestas celulares y humorales que el individuo ha desarrollado a lo largo de su vida.
INMUNIDAD INNATA
BARRERAS ANATÓMICAS
Los mecanismos de barrera son responsables de la primera línea de defensa contra los agentes patógenos y el más importante es la piel1. La piel representa una superficie difícil de penetrar y provee una excelente defensa contra la infección. Ello explica porque existe un incremento en la incidencia de infecciones locales y sistémicas cuando esta barrera esta comprometida, por ejemplo en los casos de enfermedades propias de la piel o quemaduras.
Los patógenos pueden acceder al organismo a través de otras áreas anatómicas. Los tractos respiratorio, gastrointestinal y genitourinario son sitios en los cuales las membranas especializadas de las mucosas juegan un importante papel como barrera para prevenir la infección; hay que resaltar que no solo mecanismos físicos y químicos están envueltos. Existen factores inmunológicos esenciales que defienden contra la infección en estos tejidos. La inmunoglobulina A (IgA) es un anticuerpo secretado por las células de la mucosa y que se fija al patógeno y a sus toxinas para neutralizarlos y desactivarlos antes de que los mismos sean capaces de invadir.
CÉLULAS Y PROTEINAS
Las células y proteínas del sistema innato representan una línea de defensa que está presente desde el nacimiento. A diferencia del sistema adquirido su respuesta no es alterada por subsecuentes invasiones ya que ellas no tienen memoria. Si una barreara anatómica se rompe los patógenos pueden penetrar el tejido subyacente. A este nivel el sistema innato provee defensas no específicas que previenen el crecimiento bacteriano y la consecuente infección. Este sistema se enfrenta a la invasión a través de respuestas celulares y proteínas del complemento, esto último se discutirá al final de este tutorial.
La respuesta celular1,2,3 se basa en los leucocitos que pueden ser granulocitos (neutrófilos, eosinófilos y basófilos) o agranulocitos (monocitos y linfocitos). La meta del sistema innato es destruir los patógenos para prevenir la diseminación de la infección. Un arma importante en este sentido es la fagocitosis (figura 1), allí se describe el proceso por el cual la célula engloba el material extraño y lo elimina. Los macrófagos y neutrófilos son la primera línea fagocítica del sistema innato pero otras células también juegan algún papel.
Los macrófagos son las primeras células fagocíticas y son una de las dos líneas celulares que maduran desde los monocitos. Los monocitos también se diferencian en células dendríticas. Estos tienen actividad fagocítica limitada pero juegan un papel crucial en conjugar el sistema inmune innato con el adquirido al cumplir el papel de célula presentadora de antígenos. La importancia de esto será discutirá mas adelante
Para que la fagocitosis tenga lugar, el patógeno en primer lugar debe ser reconocido como extraño. Esto ocurre en virtud de interacciones entre receptores especializados de superficie de la célula del macrófago con patrones moleculares asociadas a patógenos, expresados por los microbios. Un ejemplo de esta interacción está dado por el receptor toll-like (TLR) del macrófago que reconoce a los lipopolisacáridos de las bacterias gramnegativas.
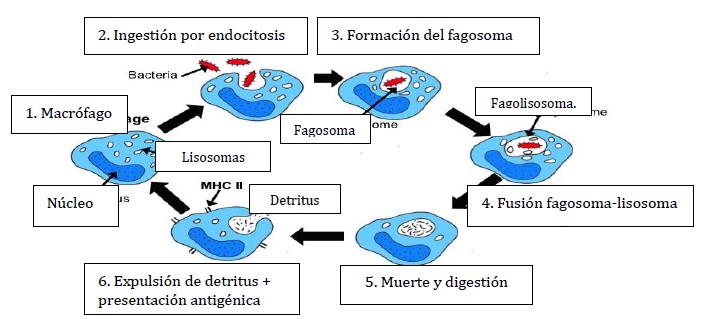
Figura 1: Interacción del microbio con el macrófago estimulando la fagocitosis, El fagosoma se fusiona con los lisosomas del macrófago los cuales acidifican y digieren al microbio. El macrófago presenta partículas microbianas en su superficie en asociación con el antígeno mayor de histocompatibilidad clase II (MHC II) los cuales permiten la activación de otras células inmunes.3
Una segunda clase de fagocitos son los leucocitos polimorfonucleares, más comúnmente conocidos como neutrófilos. Estas células son reclutadas tempranamente durante la infección por los macrófagos a través de la liberación de citokinas proinflamatorias como la Interleukina 1β (IL-1β), IL-6, IL 12 e IL-18. Así se incrementa el número de células fagocitarias a nivel tisular para ayudar a destruir al enemigo patógeno. En adición a las citokinas mencionadas arriba, los macrófagos liberan factor de necrosis tumoral ∞ (TNF-∞), prostaglandinas, leucotrienos y factor activador plaquetario que inducen los cambios vistos en la inflamación aguda tales como incremento en la permeabilidad vascular, coagulación e intensidad del dolor. El objetivo de esta respuesta es prevenir la diseminación de la infección por vía hemática y reclutar futuras células
inmunes.
No todos los invasores son extracelulares. Los virus infectan las células del huésped y pueden evadir la fagocitosis de los macrófagos. El sistema inmune innato es capaz de combatir las infecciones intracelulares mediante otra línea celular, los natural killer o células NK. Gracias a las citokinas proinflamatorias las NK son reclutadas en el sitio de infección y logran la detección de las células infectadas por medio del reconocimiento de una alteración o ausencia del complejo mayor de histocompatibilidad (MHC) clase 1. (Figura 2)
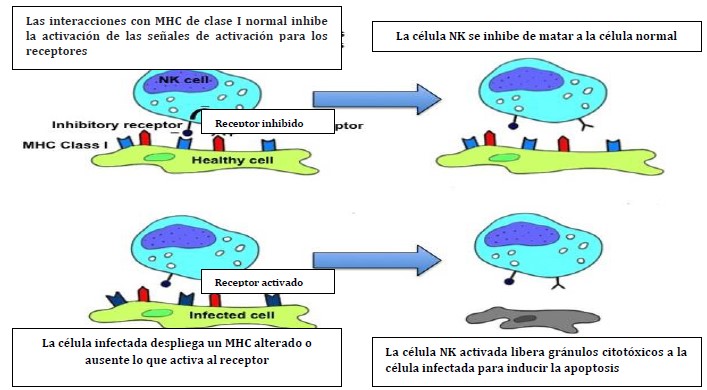
Figura 2. Arriba: Células sanas muestran un complejo MHC normal sin activar las células NK. Abajo: Células infectadas despliegan un MHC anormal o alterado generando un gatillo para la activación de las células NK.1
INMUNIDAD ADQUIRIDA
La inmunidad adquirida describe respuestas a la infección que han sido aprendidas por el sistema inmune con el tiempo. La inmunidad innata no siempre puede controlar la infección por si sola y entonces trabaja al unísono con las células y proteínas altamente especializadas del sistema inmune adquirido.
Después de la primera exposición a un patógeno, la respuesta adquirida toma tiempo en desarrollarse. No obstante debido a la memoria de este sistema, las infecciones subsecuentes producidas por el mismo agente generan una respuesta rápida. La administración de vacunas individuales representa la mayor ventaja de este tipo de respuesta.
El sistema adquirido es clasificado en 2 componentes: componente humoral representado por las células B y sus proteínas y componente celular representado por los linfocitos T (células T). Ambas líneas celulares son generadas desde células madres del sistema hematopoyético en la médula ósea pero son subclasificadas de acuerdo a su sitio de diferenciación: Las células T se desarrollan en el timo y las B en la médula ósea.
Las células T y B expresan en su superficie celular receptores altamente específicos. El genoma mamífero tiene códigos para millones de receptores diferentes de las células T y B. Durante su desarrollo las células son seleccionadas por su capacidad para reconocer estos receptores de antígenos específicos. Es importante destacar que cualquier célula que responde fuertemente a las propias proteínas es eliminada para evitar un ataque inmune contra las propias células del organismo, este es un proceso que puede estar defectuoso en las enfermedades autoinmunes. Las células que responden fuertemente a los péptidos no propios entonces permanecen inertes en los tejidos linfoides secundarios del bazo, ganglios linfáticos o el tejido linfoide asociado a mucosas (MALT) hasta su activación producida cualquier futura infección.
CELULAS T
Las células T juegan un papel en la defensa contra los patógenos tanto intracelulares (virus) como extracelulares (bacterias). Estas células son seleccionadas por la capacidad de su receptor T (TCR) de reconocer al MHC asociados a péptidos no reconocidos por el organismo.
MHC se encuentran desplegados en todas las células de los mamíferos y son específicos para cada individuo. MHC clase I es expresado por todas las células nucleadas mientras que el MHC clase II es expresado solo por células del sistema inmune, predominantemente por células presentadoras de antígenos. Es la interacción del receptor de células T con el MHC clase I o clase II la que da lugar a la clasificación de estas células; CD4 + y CD8 +. Las células T inmaduras expresan co-receptores en las células CD8+ y CD4+ que tienen afinidad por el MHC clase I y clase II, respectivamente. Durante su desarrollo en el timo, las células T están expuestas al péptido antigénico asociado al MHC clase I o clase II. Si los TCR se asocian fuertemente con MHC de clase I, el timocito conservará entonces su receptor CD8+ pero pierden su receptor CD4+. Lo contrario ocurre si el TCR tiene afinidad por el MHC de clase II. Tras el desarrollo en el timo, las células T ingenuas residen dentro del tejido linfoide secundario, ya sea como CD8 + o células CD4 + esperando a ser activados.
La activación de células T inertes ocurre durante un proceso llamado “presentación antigénica”. Las células dendríticas y los macrófagos envueltos en la respuesta innata migran al tejido linfoide donde presentan el antígeno asociado con el MHC, por esto, ellas son llamadas células presentadoras de antígenos (CPAs). Las células T CD8+ son activadas si su péptido específico es presentado al MHC clase I y las CD4+ se activan si es presentado al MHC clase II. La activación de las células T CD8+ origina células T citotóxicas. Como el MHC clase I está presente en todas las células nucleadas, si una célula es infectada por un virus ella desplegará un péptido no reconocido al MHC. Las células CD8+ especifica para este antígeno destruirá estas células por liberación de vesículas citoplásmicas a la célula infectada, estas vesículas contienen caspasas que inician la muerte celular programada o apoptosis.
Hay 4 tipos de células T CD4+, a saber; T-cooperadora 1 (Th1), T-cooperadora 2 (Th2), T-cooperadora 17 (Th17) y Treguladora. La formación de cada subpoblación depende del entorno de citokinas en el cual el antígeno es presentado a la célula T y cada subpoblación cumple un rol diferente en la inflamación. La tabla 1 muestra las diferencias en los diversos tipos de células CD4+. Es importante notar las acciones inhibitorias de cada clase sobre la otra lo que permite una más efectiva defensa ante los patógenos.
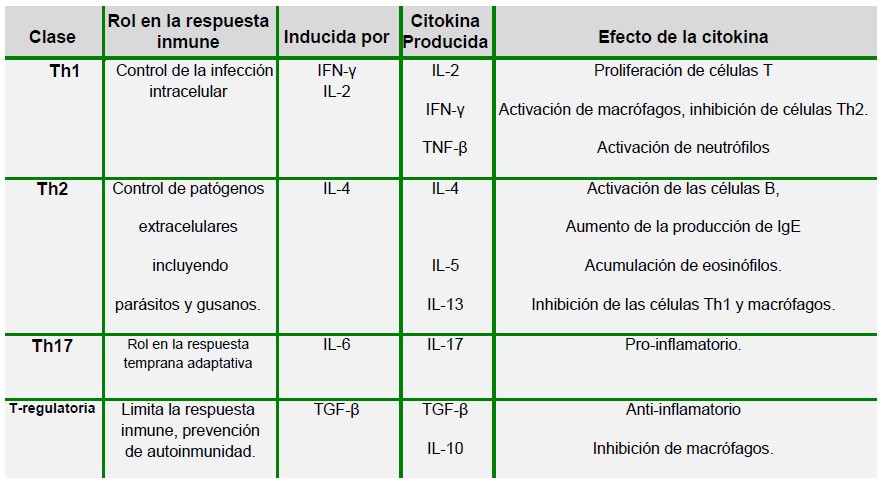
Tabla 1: Subclasificación de las células T CD4+. Interferón (IFN), Factor de Crecimiento Tumoral (TGF) Factor de Necrosis Tumoral (TNF).
Las células NKT son una subclase de células T independiente de la clasificación. Estas células son capaces de reconocer extraño y liberar IL-4 y IFN-Y aunque parecen favorecer la respuesta tipo Th2. No deben ser confundidas con las células NK de la respuesta innata.
Células B 1,2,4.
Las células B están envueltas en la respuesta humoral inmune, ellas median estas respuestas a través de la producción de immunoglobulinas (Ig) o anticuerpos cuyo papel es proteger contra patógenos extracelulares.
Las células B presentan un receptor altamente específico (BCR) con afinidad por un solo antígeno. Este receptor está unido a la membrana de la molécula de inmunoglobulina que es secretada por la célula B cuando se activa. Cuando el BCR interactúa con su antígeno, se internaliza el complejo antígeno-receptor, procesa el antígeno para luego mostrar péptidos antigénicos en asociación con el MHC clase II en su superficie celular. De esta manera las células B actúan como CPAs de células T auxiliares que han sido activados por el mismo patógeno. Las células T entonces liberan las citokinas IL-2, IL-4, IL-10 e IL-13 que inducen la activación de células B. En raras ocasiones, algunas células B se pueden activar sin la ayuda de células T. Este fenómeno sólo se produce en la presencia de antígenos extraños que pueden inducir directamente vías de señalización celular en la interacción con el BCR.
Consecutivamente a su activación las células B proliferan y se diferencian en un gran número de idénticas células plasmáticas y células memoria en un proceso conocido como expansión clonal. Las células plasmáticas son responsables de la secreción de immunoglobulinas mientras que las células memoria entran en un estado de reposo para una rápida y vigorosa respuesta ante una subsecuente infección producida por ese patógeno particular.
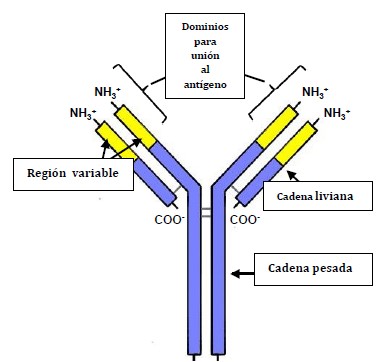
Figura 3: Estructura de immunoglobulina5
Inmunoglobulinas1,2,4
La inmunoglobulina es un complejo proteínico que consiste en dos cadenas pesadas idénticas y dos cadenas livianas también idénticas. En el extremo amino terminal de estas moléculas hay un sitio de unión altamente específico para los antígenos. Esta porción es llamada región variable, la porción del extremo carboxilo terminal es llamada región constante y es la responsable de la interacción con los receptores de anticuerpos en los neutrófilos, macrófagos y mastocitos siendo capaz además de inducir la activación de la cascada del complemento. Las moléculas de immunoglobulinas pueden ser clasificadas en 5 isoformas dependiendo del tipo de cadena pesada que las conformen (tabla 2).

Tabla 2: Isoformas de las inmunoglobulinas4
Como se muestra en la tabla 2 las inmunoglobulinas cumplen diversos roles en la defensa contra las infecciones intra y extracelulares. El rol de la Ig E en la activación de los mastocitos, basófilos y eosinófilos no debe pasarse por alto.
La Ig E es producida por células B activadas. Aunque es la inmunoglobulina menos abundante es vital para combatir la infección parasitaria. Los mastocitos, basófilos y eosinófilos son granulocitos que contienen mediadores inflamatorios que incluyen histamina, leucotrienos y TNF. Estas células expresan receptores Fc (FcR) específicos para Ig E. Cuando el complejo Ig E: FcR es ligado al antígeno las células se degranulan causando inflamación y ayudando a la destrucción del parásito. Este proceso también está presente y contribuye a las reacciones alérgicas y de hipersensibilidad.
Mientras que la respuesta celular es crítica para la efectividad de la respuesta innata, un mecanismo dependiente de una segunda proteína conocido como el sistema de complemento tiene un rol igualmente importante.
Sistema de Complemento1,2,4
El hígado produce un número de proteínas del complemento que están implicados tanto en la inmunidad innata y adquirida. Aquí, la cadena de acontecimientos se activa por la invasión microbiana y el proceso resultante se conoce como la cascada del complemento. Hay dos formas principales por las que el sistema del complemento destruye los microbios a saber opsonización y la cascada del complemento.
La opsonización es el proceso a través del cual los patógenos son marcados por las proteínas del complemento, para luego inducir la fagocitosis por macrófagos que expresan receptores específicos para dichas opsoninas. La fijación de las proteínas del complemento también activa la cascada del complemento el cual es un proceso más complejo de activación de proteínas que induce directamente la muerte microbiana a través de la destrucción de la membrana del patógeno.
La cascada del complemento puede ser activada por tres vías: clásica, lectina y alterna. Cada vía inicialmente implica diferentes proteínas del sistema, pero todas llevan a la hidrólisis de la proteína de complemento C3 a C3a y C3b. C3a actúa como una citokina inflamatoria, mientras que C3b actúa como una opsonina o como iniciador del ataque de la membrana del patógeno.
El desarrollo exacto de la cascada del complemento es muy complejo y se escapa de los fines de este tutorial, no obstante tres fragmentos proteicos dentro de esta cascada son importantes de considerar. C3a, C4a y C5a los cuales funcionan como potentes citokinas inflamatorias causando contracción del músculo liso e incrementando la permeabilidad vascular y la degranulación de los mastocitos y de los basófilos. Estas proteínas son conocidas como anafilotoxinas debido a su capacidad para inducir reacciones anafilácticas independiente de la Ig E. La anafilaxis
será estudiada en detalle en el tutorial dos.
Resumen
El sistema inmune es una compleja red de células y proteínas capaces de proteger contra la infección. La respuesta innata es llevada a cabo mayormente por los fagocitos y las proteínas del complemento en tanto que la inmunidad adquirida se basa en un una respuesta especializada celular y humoral que involucra a las células T y B. Los dos sistemas están íntimamente interconectados y trabajan de manera sinérgica en la lucha contra los invasores patógenos.
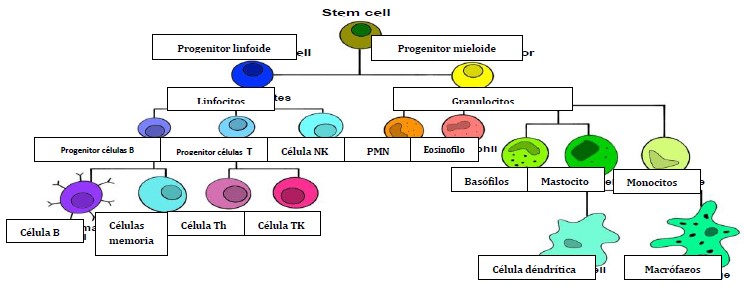
Figura 4. Células del sistema inmune.
Respuestas
- ..
a) Verdadero: Un defecto en el sistema de inmunidad mediado celularmente puede generar la autodestrucción inapropiada de las células del propio organismo pudiendo desarrollarse entonces enfermedades autoinmunes.
b) Verdadero: La activación de linfocitos B ocurre en la médula ósea y de los linfocitos T en el timo.
c) Falso: Las células plasmáticas forman parte de la respuesta humoral, no de la mediada por células.
d) Falso: Las inmunoglobulinas no participan en la respuesta inmune mediada celularmente. La IgA es una inmunoglobulina mucosal.
e) Falso: La histamina es liberada por los mastocitos cuando son activados por la IgE, lo cual no es parte del sistema inmune mediado por células. - ..
a) Verdadero: Las células B son nombradas así luego de madurar en la medula ósea (Bone). Las células T maduran en el Timo.
b) Verdadero: La forma de los receptores celulares en la superficie de las células B es similar a la inmunoglobulina que la célula produce, esto asegura que la inmunoglobulina se unirá al antígeno específico identificado por la célula B.
c) Falso: Las células NK son parte de la respuesta innata inmune.
d) Falso: Las células B son activadas principalmente por células T. Ocasionalmente algunos antígenos pueden activarlas de manera directa.
e) Falso: Cada célula B individual solo produce un solo tipo de anticuerpos. - ..
a) Falso: Las inmunoglobulinas son producidas por las células plasmáticas (células B).
b) Verdadero: La IgA es un tipo de inmunoglobulina que se encuentra en las membranas mucosas.
c) Falso: Cada inmunoglobulina consiste en dos cadenas pesadas y dos cadenas livianas.
d) Falso: La especificidad para los antígenos es dependiente de la región variable zona que involucra a ambas cadenas pesadas y livianas.
e) Falso: La IgE causa la degranulación del mastocito.
Referencias.
- Murphy K, Travers P, Walport M; Immunobiology, 7th Ed. New York, Garland Science, 2008.
- Todd I, Spickett G; Lecture Notes Immunology, 6th Ed. Oxford, Wiley-Blackwell, 2010
- Todar, P. (2012). Immune Defense against Bacterial Pathogens: Innate Immunity. Available: http://textbookofbacteriology.net/innate.html Last accessed 2nd July 2015.
- Male, D; Immunology An Illustrated Outline, 3rd Ed. London, Mosby-Wolfe, 1998.
- Biochemistry Department, University of Otago. (1999). Biochemistry 111. Available: http://biocadmin.otago.ac.nz/fmi/xsl/bioc/learnbitslecture.xsl?-db=BIOCweb.fp7&-lay=Lectures&- recid=3970&-find Last accessed 2nd July 2015.
This work by WFSA is licensed under a Creative Commons Attribution-NonCommercial-NoDerivitives 4.0 International License. To view this license, visit https://creativecommons.org/licenses/by-nc-nd/4.0/