Basic Sciences

Introduction
The neuromuscular junction is made up of a motor neurone and a motor endplate with a synaptic cleft or junctional gap dividing them. It is critical in the production of skeletal muscle contraction.
The muscle relaxants used by anaesthetists in daily practice and a number of diseases have their effect at the neuromuscular junction and an understanding of its structure and physiology is therefore important.
Before reading the tutorial, the following questions will establish the knowledge you already have on this topic.
Questions
- What is the structure of the neuromuscular junction?
- How is acetylcholine synthesised, stored and released?
- What is the structure of the acetylcholine receptor and how does it work?
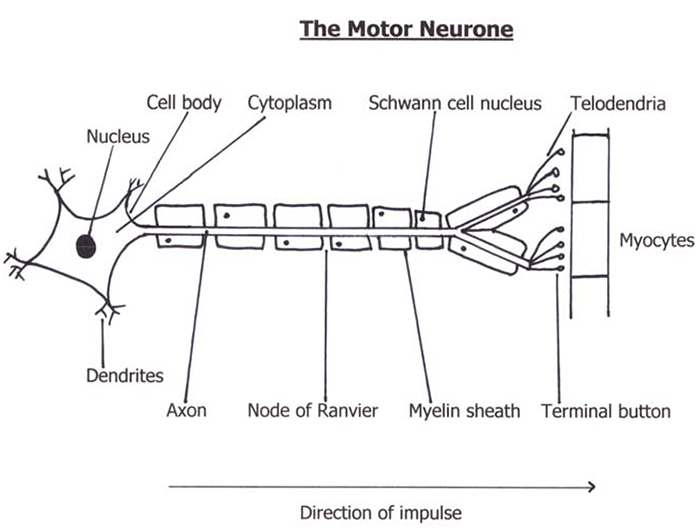
The Motor Neurone
Motor neurones are the nerves that control skeletal muscle activity. They originate in the ventral horn of the spinal cord and travel up to a metre to the muscles they supply. The cell body of a neurone is at its proximal end and information travels from here down the axon. Axons are 10-20μm in diameter and surrounded by a myelin sheath produced by Schwann cells. This acts as an insulator to speed up nerve conduction. The myelin sheath is interrupted by nodes of Ranvier between which the action potential jumps causing rapid conduction of the nerve impulse (saltatory conduction).
Each motor neurone connects to several skeletal muscle fibres to form a motor unit. The number of muscle fibres within the motor unit varies enormously, from a few, for fine motor control (e.g. the muscles of the eye), to several thousand for coarse actions (e.g. the thigh muscles). There is however only one neuromuscular junction on each skeletal muscle fibre, any others are eliminated during development.
As the motor neurone enters a muscle, the axon divides into telodendria, the ends of which, the terminal buttons, synapse with the motor endplate. The two are separated by approximately 20nm, the junctional gap or synaptic cleft. It is here that release of the neurotransmitter acetylcholine occurs with consequent binding to the receptors on the motor endplate.
The Motor Endplate
The motor endplate is a highly specialised region of the sarcolemma of a muscle fibre. It is oval in shape and covers an area of about 3000μm2 . Its surface is deeply folded with multiple crests and secondary clefts. The nicotinic acetylcholine receptors are located on the crests of the folds in excessive numbers (1-10 million) and concentration (10,000-20,000 μm2) to ensure the success of this effector system. The clefts of the motor endplate contain acetylcholinesterase.
The area of muscle around the motor endplate is the peri-junctional zone. It is here that the potential developed at the endplate is converted to an action potential that propagates through the muscle to initiate contraction. The peri-junctional zone has an enhanced ability to produce a wave of depolarisation to the muscle from that produced by the post-synaptic receptors.
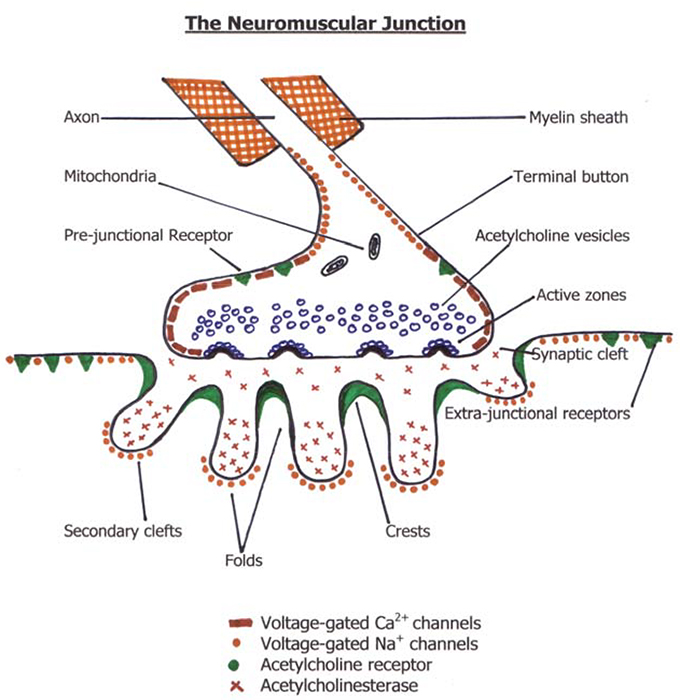
Acetylcholine synthesis, storage and release
Acetylcholine is synthesised from choline and acetyl-coenzyme A (acetyl-coA) in the terminal axoplasm of motor neurones and is catalysed by the enzyme choline acetlytransferase. Acetyl-coA is synthesised from pyruvate in the mitochondria in the axon terminals. Approximately 50% of the choline is extracted from extracellular fluid by a sodium dependant active transport system, the other 50% is from acetylcholine breakdown at the neuromuscular junction. Overall, the majority of the choline originates from the diet with hepatic synthesis only accounting for a small proportion.
Choline acetyltransferase is produced on the ribosomes in the cell body of the motor neurone from where it is transported distally by axoplasmic flow to the terminal button and can be found in high concentrations. The activity of choline acetyltransferase is inhibited by acetylcholine and increased by nerve stimulation.
Once synthesised the molecules of acetylcholine are stored in vesicles within the terminal button, each vesicle containing approximately 10,000 molecules of acetylcholine. These vesicles are loaded with acetylcholine via a magnesium dependent active transport system in exchange for a hydrogen ion. The vesicles then become part of one of three pools or stores, each varying in their availability ability for release. About 1% are immediately releasable, about 80% are readily releasable and the remainder form the stationary store. The exact proportions may vary depending on the level of demand or nerve stimulation.
The release of acetylcholine into the synaptic cleft may be spontaneous or in response to a nerve impulse. Spontaneous release of single vesicles of acetylcholine occurs randomly and results in miniature endplate potentials of 0.5-1mV, the function of which is unknown. With the arrival of a nerve impulse, large numbers of P-type calcium channels in the terminal membrane of the nerve open, allowing calcium to enter the cell. The combination of depolarisation of the presynaptic terminal and influx of calcium triggers 100-300 vesicles to fuse with the presynaptic membrane at specific release sites opposite the junctional folds and release acetylcholine into the synaptic cleft (exocytosis). This causes a brief depolarisation in the muscle that triggers a muscle action potential (see below). The depleted vesicles are rapidly replaced with vesicles from the readily releasable store and the empty vesicles are recycled. At rest the free calcium concentration is kept below 10 –6M by a low membrane permeability to calcium, an active sodium/calcium exchange pump and mitochondrial sequestration.
Acetylcholine Receptors
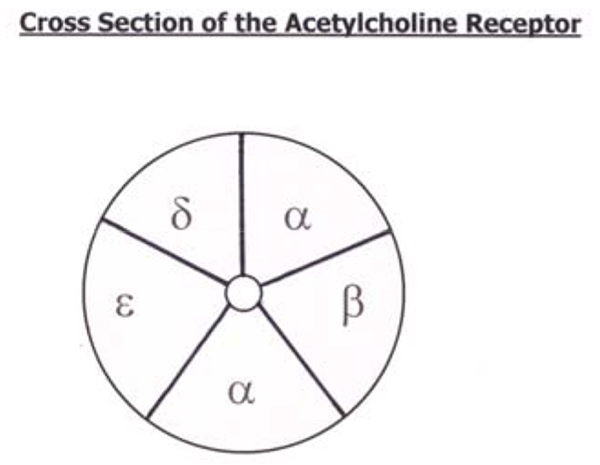
The post-junctional membrane receptors of the motor endplate are nicotinic acetylcholine receptors. There are on average 50 million acetylcholine receptors on a normal endplate, situated on the crests of the junctional folds. Each nicotinic receptor is a protein comprised of five polypeptide subunits that form a ring structure around a central, funnel-shaped pore (the ion channel). The mature adult receptor has two identical α (alpha) subunits, one β (beta), one δ (delta) and one ε (epsilon) subunit. In the foetus a γ (gamma) subunit replaces the ε. These different proteins are each coded by a different gene and synthesised within the muscle cells. The whole receptor spans the muscle cell membrane projecting predominantly extracellularly.
Acetylcholine molecules bind to specific sites on the α subunits and when both are occupied a conformational change occurs, opening the ion channel for just 1 msec. The channel allows movement of all cations, however it is the movement of sodium that predominates in terms of both quantity and effect. This causes depolarisation, the cell becomes less negative compared with the extracellular surroundings. When a threshold of –50mV is achieved (from a resting potential of –80mV), voltage- gated sodium channels open, thereby increasing the rate of depolarisation and resulting in an end plate potential (EPP) of 50-100mV. This in turn triggers the muscle action potential that results in muscle contraction. By this method the receptor acts as a powerful amplifier and a switch (acetylcholine receptors are not refractory).
In addition to the post-junctional receptors on the motor endplate, acetylcholine receptors can also be found outside the neuromuscular junction and are called extra-junctional receptors, or on the pre-terminal bulb and are called pre-junctional receptors. The extra-junctional receptors can be present anywhere on the muscle membrane usually in extremely small numbers, though they are found in their greatest concentration around the endplate in the peri-junctional zone. Denervation injuries and burns are associated with large increases in the number of extra-junctional receptors on the muscle membrane. The extra- junctional receptors have the structure of immature foetal receptors (ε subunit replaced by a γ subunit). This affects the physiology and pharmacology of the receptor with increased sensitivity to depolarising muscle relaxants and reduced sensitivity to non-depolarising muscle relaxants.
Pre-junctional receptors on the terminal bulb have a positive feedback role. In very active neuromuscular junctions acetylcholine binds to these receptors and causes an increase in transmitter production via a second messenger system. These receptors may also play a role in the “fade” seen in non-depolarising muscle relaxant blockade by inhibiting replenishment of acetylcholine.
Acetylcholinesterase
In order for the acetylcholine receptor to function effectively as a “switch” it is essential that acetylcholine is removed rapidly from the junctional gap or synaptic cleft. This is achieved by hydrolysis of acetylcholine to choline and acetate in a reaction catalysed by the enzyme acetylcholinesterase (AChE). The active site in the AchE molecule has two distinct regions, an ionic site possessing a glutamate residue and an esteratic site containing a serine residue. Hydrolysis occurs with transfer of the acetyl group to the serine group resulting in an acetylated molecule of the enzyme and free choline. The acetylated serine group then undergoes rapid, spontaneous hydrolysis to form acetate and enzyme ready to repeat the process. The speed at which this occurs can be gauged by the fact that approximately 10,000 molecules of acetylcholine can be hydrolysed per second by a single site.
This enzyme is secreted by the muscle cell but remains attached to it by thin collagen threads linking it to the basement membrane. Acetylcholinesterase is found in the junctional gap and the clefts of the post- synaptic folds and breaks down acetylcholine within 1 msec of being released. Therefore the inward current through the acetylcholine receptor is transient and followed by rapid repolarisation to the resting state.
Summary
- The transmitter at the neuromuscular juntion is acetylcholine, which is synthesised from acetyl- coenzyme A and choline, stored in vesicles and released in response to a nerve stimulus.
- The acetylcholine receptor is made of 5 subunits and acts as a cation channel causing depolarisation at the motor end plate.
- The acetylcholine receptor acts as an amplifier and a switch.
- Acetylcholine is broken down to choline and acetate, catalysed by acetylcholinesterase.
- The neuromuscular junction is so vital to life that everything is done to excess, excess transmitter release, excess receptor numbers and an excessive post-synaptic potential to ensure transmission of the stimulus and skeletal muscle contraction.
This work by WFSA is licensed under a Creative Commons Attribution-NonCommercial-NoDerivitives 4.0 International License. To view this license, visit https://creativecommons.org/licenses/by-nc-nd/4.0/