Basic Sciences

QUESTIONS
Before continuing, try to answer the following questions. The answers can be found at the end of the article.
- What percentage of total blood volume is composed of plasma?
a. 100%
b. 75%
c. 55%
d. 30%
e. 25% - What is the lifespan of a red blood cell?
a. 180 days
b. 120 days
c. 90 days
d. 12 days
e. 1 day - What factors influence oxygen affinity?
- How is carbon dioxide transported in the blood?
- What are the buffering components of blood?
INTRODUCTION
This is the first of two tutorials on blood physiology for the anaesthetist. This tutorial will outline the production and constituents of blood, as well as blood group s, gas transport and the role of blood in acid base balance The second tutorial will outline the complex processes involved in haemostasis.
Blood is the fluid constituent of the cardiovascular system, comprising both plasma and cellular components. Fifty five percent of total blood volume is composed of plasma, which in turn comprises 90% water, 8% plasma proteins, with the remaining 2% consisting of electrolytes and organic compounds. Red blood cells, white blood cells and platelets form the cellular component. Blood has many essential homeostatic functions including the transport of oxygen and carbon dioxide, nutrients and waste products, immune function, buffering and haemostasis. As blood has such an important role in the maintenance of homeostasis, the anaesthetist needs a good knowledge of its constituent s and physiology.
HAEMOPOIESIS
Sites of Production
The main sites of haemopoiesis or blood cell production differ during the various stages of human development. Within the first few weeks of the gestational period haemopoietic cells appear in the yolk sac. From approximately six weeks onwards the liver becomes the main site of blood cell production, with the spleen and lymph nodes playing a smaller contributory role. It is during the last month of gestation that the red marrow becomes the main site of blood cell production. Red bone marrow is found within all bones up until the age of five years old, when it is gradually replaced by yellow bone marrow which comprises mainly fat. From the age of twenty years onwards the production of most blood cells is confined to the vertebrae, sternum, ribs and ilia.
Pluripotential Haemopoietic Stem Cells
These cells are found in the bone marrow and it is from these cells that all types of blood cells are derived. They give rise to daughter cells, which in turn differentiate into progenitor stem cells from which precursor cells for each different blood cell type are derived. It is from these precursor cells that mature blood cells develop. (Table. 1)
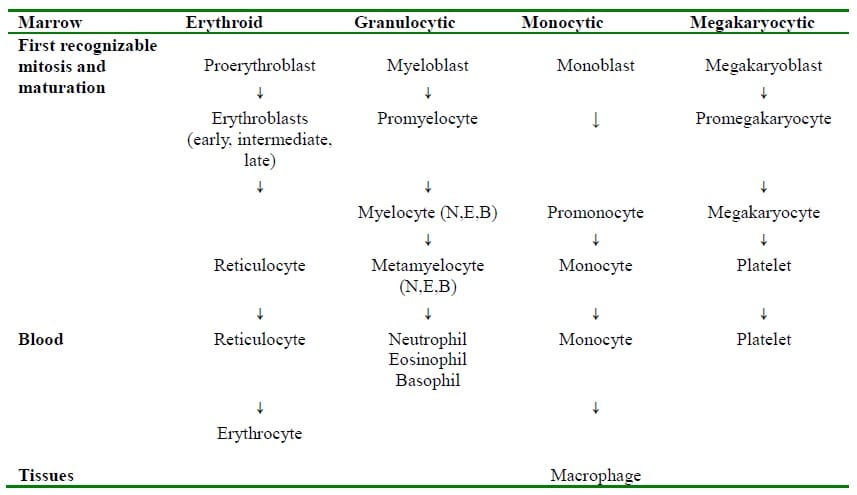
Table 1. Bone marrow production lines (Adapted from Lecture Notes Human Physiology, 5th edition, 2009)
Erythropoiesis
The production of red blood cells or erythropoiesis from the precursor the proerythroblast to the formation of reticulocytes and their release into the circulation takes about seven days. This process is under the control of erythropoietin , which is a glycoprotein hormone synthesized mainly in the kidney, with 10 15% being produced by the liver. In the fetus, however, the liver is the major site of erythropoietin production. Erythropoietin is produced by the peritubular capillaries of the kidney in response to local hypoxia. This local decrease in oxygen delivery may be due either to anaemia or systemic hypoxaemia . Once erythropoietin is released it acts on immature erythroid cells in the bone marrow by binding to a specific receptor found in their cell membrane, causing them to differentiate and proliferate into mature red blood cells. The rate of red cell production is closely balanced by the rate of destruction.
The Red Blood Cell
The mature red blood cell (RBC) is a biconcave disc approximately 7-8 micrometre in diameter with out a nucleus. The size and shape not only increase the surface area of the RBC, thereby facilitating gas exchange, but also increase flexibility , allowing passage through narrow capillaries. The lifespan of a RBC is approximately 120 days and is removed from the circulation by macrophages found in the bone marrow, spleen and liver.
Haemoglobin
Haemoglobin is a large protein found within red blood cells. Synthesis is initiated in the bone marrow by erythroblasts and is completed once the cells become reticulocytes. It is composed of four subunits with each subunit consisting of a polypeptide with a haem moiety. There are two pairs of polypeptide chains I n normal adult haemoglobin (HbA) these are named alpha and beta chains. The four chains comprise the globin part of the molecule. The haem moiety consists of a porphyrin ring with a central iron atom. Oxygen binds to ea ch iron atom to form oxy-haemoglobin, enabling each haemoglobin molecule to carry four molecules of oxygen.
Iron Metabolism
Iron is an essential component for haemoglobin production. Most of the iron for this process comes from the recycling of other iron stores with only 1 2mg per day being absorbed from the diet. Absorption of iron occurs in the duodenum and is tightly controlled by duodenal enterocytes. Iron is absorbed in three forms: Ferrous ion (Fe 2+ ), Ferric ion ( Fe 3+ ) and haem. Within the en terocytes Fe 2+ forms ferritin where it is stored as Fe 3 + by binding to apoferritin. It is then transported across the basolateral membrane by ferroportin. Ferroportin is coupled to feroxidase which converts the ferrousion to its ferric form. Only Fe 3+ c an pass freely from the enterocyte into the bloodstream. Once in the bloodstream Fe 3+ is carried by transferrin to its target cell. The iron is released by endocytosis of the transferrin complex and after removal of the iron the transferrin molecule return s to the circulation. The levels of transferrin are directly affected by the iron stores, with adequate stores resulting in lower levels of transferrin. This in turn results in decreased transfer of iron across the duodenal mucosa.
White Blood Cells
The white blood cells are responsible for the body’s defence mechanisms. There are two main groups of white cells, namely phagocytes and lymphocytes. Phagocytes are cells that engulf and destroy foreign material. They comprise monocytes (the precursors of macrophages ) and granulocytes . Granulocytes consist of three cell types: neutrophils , eosinophils and basophils. Lymphocytes are the effector cells of the immune system and comprise B and T lymphocytes.
B Lymphocytes
B Lymphocytecells mature in the bone marrow , hence are called B cells. They comprise 15% of the total lymphocyte population and pass between the spleen/lymph nodes and the circulation. Their role is to mature into antibody producing plasma cells after encountering a specific antigen. An antigen is any molecule that can bind to an antibody. An antibody or immunoglobulin is a protein that binds to an antigen, priming it for phagocytosis. Once an immature B cell encounters an antigen that matches its membrane bound immunoglobulin it ingests it. Major Histocompatibility Complex (MHC) II molecules found on the cell surface of the B cell then present the fragments of the encountered antigen. In order to mature into an antibody producing plasma cell this complex must bind with its specific matching type 2 T helper (TH 2) cell, initiating the release of cytokines from the TH 2 cell, thereby inducing the B cell to complete its maturation (Fig. 1)
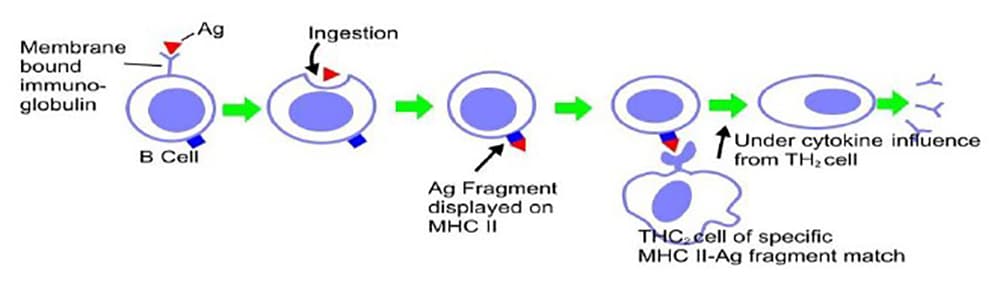
Figure 1. The development of plasma cells in response to antigen binding on the B cell surface
T Lymphocytes
These cells differentiate in the thymus, hence are named T cells. Similar to B cells they also pass between the circulation and the spleen/lymph nodes. Macrophages and dendritic cells present antigens to T cells, thereby activating them to produce cytokines, which in turn results in the differentiation and proliferation of different classes of effector T cells. There are three classes of effector T cells: CD8 (cytotoxic cells) and two types of CD4 T helper cells ; Type 1 (TH 1) and Type 2 (TH 2). Cytotoxic cells kill target cells that display fragments of viral pathogens bound to MHC class I molecules on their cell surface. TH 1 cells destroy intracellular organisms by activating macrophages, whilst TH 2 cells as described above initiate the maturation of B cells into antibody producing plasma cells. (Fig. 2)
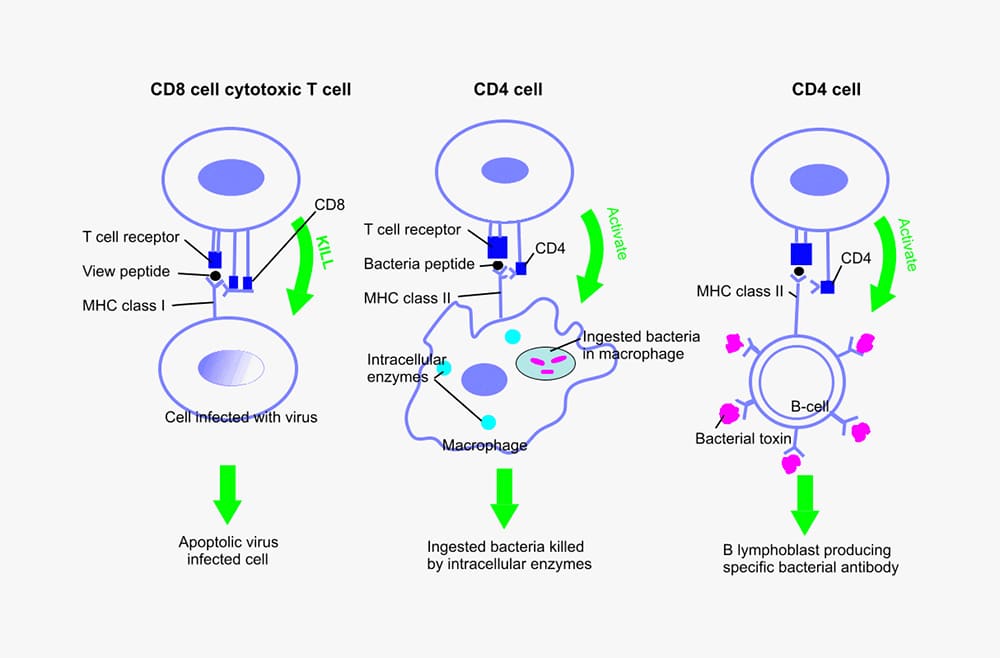
Figure 2. T cell classes and activation in response to pathogen
BLOOD GROUPS
ABO SYSTEM
Blood groups are genetically determined by the presence of the antigens found on the membranes of red blood cells. Agglutination, a process whereby cells clump together, occurs when red cells with a specific antigen encounter its corresponding antibody. In vivo agglutination results in either intravascular or extravascular haemolysis. The ABO blood group system consists of four main groups and these are determined by the presence or absence of the antigens, A and B. The presence of antigen A or antigen B gives rise to Group A or Group B, the presence of both antigens gives rise to Group AB and the absence of both antigens gives rise to Group O. Antibodies to these antigens can be either naturally occurring or as a result of an immune response. Immune antibodies arise when an individual is exposed to foreign red cell antigens through either transfusion or passage of red cells across the placenta during pregnancy.
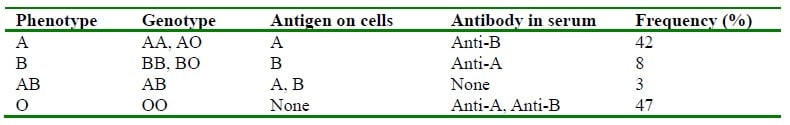
Table 2. ABO System (Reproduced from Lecture Notes Human Physiology, 5th edition, 2009)
RHESUS BLOOD GROUP SYSTEM
The Rhesus system comprises five main antigens, namely C, c, D, E and e. The term rhesus-positive usually refers to those individuals who express the D antigen on their red blood cells and rhesusnegative for those whose red cells do not express this antigen. Antibodies to these rhesus antigens occur very rarely in nature, although there are some forms of naturally occurring anti-E. The production of immune antibodies, most commonly anti-D, occurs after sensitisation by pregnancy or transfusion. It is for this reason that anti-D is injected into a rhesus-negative mother after transplacental passage of fetal blood into the maternal circulation. This destroys any fetal rhesus positive red blood cells before the maternal immune system can respond.
OXYGEN TRANSPORT
Ninety-nine percent of oxygen transported in the blood is bound to haemoglobin and 1% is dissolved in the plasma at normal atmospheric pressure.
HAEMOGLOBIN
Each molecule of haemoglobin can bind four molecules of oxygen reversibly with the ferrous iron atom of the haem moiety, to form oxyhaemoglobin. As each molecule of oxygen is unloaded the beta chains of the haemoglobin protein move apart, permitting the entry of the red cell metabolite 2,3- diphosphogylcerate (2,3 DPG). This reduces the affinity of haemoglobin for oxygen and facilitates off loading in an oxygen poor environment. Conversely the affinity of haemoglobin for oxygen is increased as each oxygen molecule is bound, so promoting uptake in an oxygen rich environment. It is this cooperative property that gives rise to the sigmoid – shaped oxygen dissociation curve (ODC). (Fig 3)
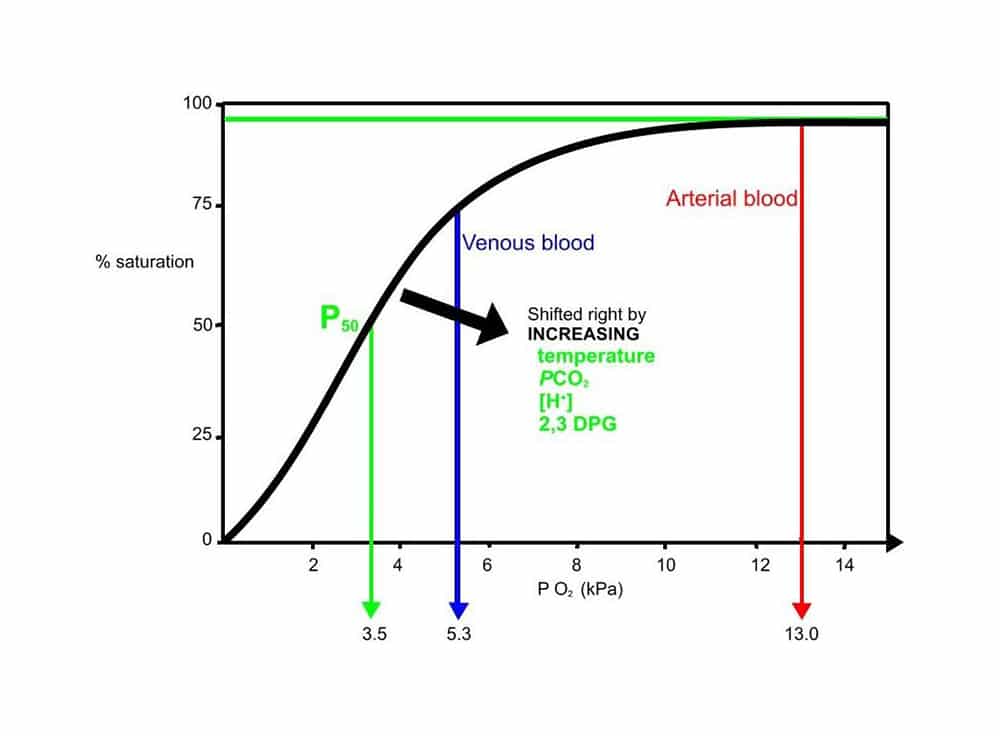
Figure 3. Oxygen haemoglobin dissociation curve
FACTORS ALTERING OXYGEN AFFINITY
A number of factors alter the position of the oxygen dissociation curve. An increase in hydrogen ions, PCO2, temperature and red cell 2,3-DPG levels shifts the curve to the right and leads to a decrease in oxygen affinity.
DISSOLVED OXYGEN
In accordance with Henry’s law the amount of oxygen dissolved in the plasma is proportional to its partial pressure. It is this dissolved fraction that diffuses into the tissues.
CARBON DIOXIDE CARRIAGE
Carbon dioxide produced by the metabolically active tissues diffuses down its concentration gradient into the blood. It is carried in three forms: dissolved CO2, as bicarbonate and as carbamino compounds. The combination of CO2 and water is catalysed by carbonic anhydrase found in red blood cells to form carbonic acid, which then undergoes fast degradation into bicarbonate and protons. The protons are taken up by haemoglobin while bicarbonate diffuses out of the red blood cells into plasma and is replaced by chloride ions to maintain electro-neutrality. Carbamino compounds are formed from CO2 reacting with the terminal amino groups of proteins and the amino groups in the side chains of arginine and lysine. Haemoglobin is the most important protein in the formation of carbamino compounds as there is more haemoglobin present and each molecule has four N-terminal groups. The percentage contributions of the three forms of carriage to total carriage and arterio-venous (a-v) difference are shown in table 3:

Table 3. CO2 Transport (Reproduced from The Physiology Viva Questions and Answers 2003)
HALDANE EFFECT
This describes the increased ability of blood to carry CO 2 as haemoglobin offloads its oxygen. Two factors are responsible for this phenomenon. Firstly, the formation of carbamino compounds is increased as oxygen is offloaded from haemoglobin. This accounts for 70% of the H aldane effect as deoxygenated haemoglobin is 3.5 times more effective in forming these compounds than oxygenated haemoglobin. Secondly, deoxygenated haemoglobin buffers the hydrogen ions produced from the dissociation of carbonic acid better than oxygenated haemoglobin, thereby improving the carriage of CO 2 as bicarbonate. (Fig 4)
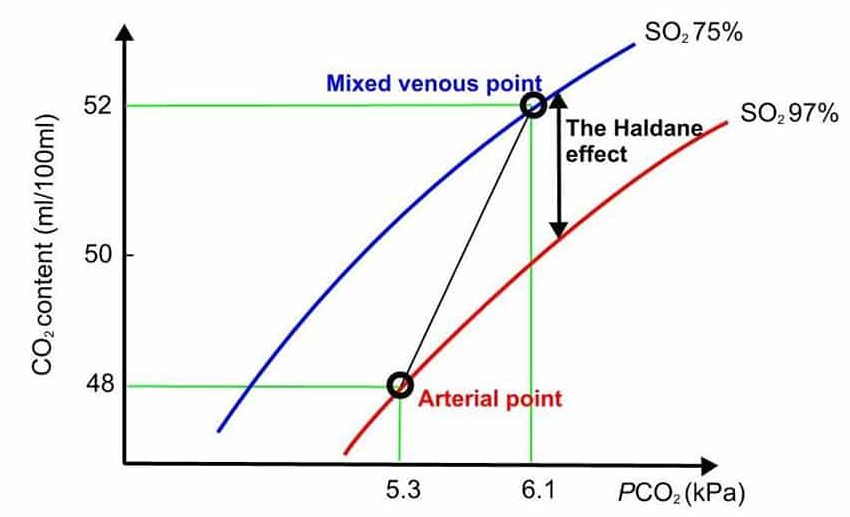
Figure 4. Carbon dioxide haemoglobin dissociation curve.
THE ROLE OF BLOOD AS A BUFFER
Buffering systems in the body minimise the change in pH (-log [H+]) that occurs when acids are added or removed. A little over half of the body’s buffering capacity is in the form of intracellular proteins and phosphates. The blood provides a little under one third of whole body buffering with two thirds of this from the carbon dioxide bicarbonate system. Haemoglobin contributes about 8% of the total body buffering capacity, with de-oxyhaemoglobin a more powerful buffer than oxyhaemoglobin. The remaining capacity is derived from plasma proteins and bicarbonate within the interstitial fluid.
ANSWERS TO QUESTIONS
- c.
- b.
- Increase s in hydrogen ion concentration (↓pH), PCO 2, temperature and 2,3 DPG all reduce the affinity of haemoglobin for oxygen and shift the ODC to the right .
- Carbon dioxide is predominantly carried in the blood as bicarbonate . The bicarbonate is derived from carbonic acid which is produced when carbon dioxide and water combine. A small quantity of carbon dioxide is dissolved in the blood and the rest is bound to proteins, predominantly haemoglobin as carbamino compounds
- Buffers reduce the change in pH when acid is added or removed from a solution . Bicarbonate is the most important buffer in the blood followed by , haemoglobin and plasma proteins.
Acknowledgements
Many thanks to Mr David Wilkinson for drawing the illustrations in figures 1, 2, 3 and 4
REFERENCES and FURTHER READING
- Brandis K. The Physiology Viva. Questions and Answers. Australia. Australia Print and Copy. 2003.
- Guyton, A; Hall J. Textbook of Medical Physiology.
- London. W.B. Saunders. 1996
- Ed: Petersen O. Lecture Notes: Human Physiology. Oxford. Blackwell Publishing. 2009
- Ed: Spoors C, Kiff K. Training in Anaesthesia the essential curriculum. Oxford. Oxford University Press. 2010
This work by WFSA is licensed under a Creative Commons Attribution-NonCommercial-NoDerivitives 4.0 International License. To view this license, visit https://creativecommons.org/licenses/by-nc-nd/4.0/