Basic Sciences

QUESTÕES
Antes de continuar, tente responder as questões abaixo. As respostas podem ser encontradas no final do artigo, juntamente com sua explicação. Por favor, responda Verdadeiro ou Falso:
- O sistema imune mediado por células:
a. Pode ser responsável por doenças autoimunes
b. Envolve a ativação de linfócitos da periferia
c. Induz a proliferação de plasmócitos
d. Resulta em um aumento da síntese de IgA
e. Acarreta uma grande liberação de histamina - A respeito das células B:
a. São assim nomeadas pois sua maduração ocorre na medula celular (bone marrow, em inglês)
b. Elas exibem receptores cuja morfologia é similar a imunoglobulina produzida por aquela célula
c. Dão origem a células NK
d. Somente são ativas por células T
e. Produzem mais de um anticorpo clonal - Imunoglobulina:
a. É secretada por células T
b. Está presente na isoforma IgA nas membranas das mucosas
c. Consiste em uma cadeia pesada e duas cadeias leves
d. A especificidade do antígeno depende da cadeia leve
e. IgG é a responsável pela degranulação dos mastócitos
Pontos Chaves
- A resposta do sistema imune inato é não especifica e destrói patógenos principalmente por fagocitose e ativação do sistema complemento.
- A resposta do sistema imune adquirido é especifica para um certo patógeno e é efetuada principalmente por células T e B, imunoglobulinas e o sistema complemento.
- Células T podem ser CD8+, citotóxicas, ou CD4+,T-helper.
- Células B secretam imunoglobulina. Esta se liga ao antígeno de patógenos e leva a célula à destruição pelo complemento ou fagócitos.
INTRODUÇÃO
A imunologia descreve a habilidade de lutar contra infecções e depende de um sistema imune funcional. Como anestesiologistas, nós desafiamos o sistema imune em nossa prática diária, seja durante a colocação de um dispositivo intravascular, intubação endotraqueal ou administração de drogas. É importante que se tenha um conhecimento dos fundamentos da imunologia, para que se possa reduzir os riscos para os nossos pacientes e entendermos porque alguns são particularmente susceptíveis a infecções.
O objetivo do presente tutorias é proporcionar uma visão geral da resposta do sistema imunológico, enquanto um Segundo tutorial focará na prática clinica.
A resposta imune pode ser dividida em dois sistemas: imunidade inata e imunidade adquirida. Como primeira linha de defesa do organismo, a imunidade inata engloba a barreira física e algumas defesas celulares. Patógenos que ultrapassam a imunidade inata, encontram a resposta do sistema imune adquirido, a qual é uma resposta sistêmica específica celular e humoral, desenvolvida durante a vida do indivíduo.
IMUNIDADE INATA
BARREIRAS ANATOMICAS
Mecanismos de barreira são responsáveis pela primeira linha de defesa contra patógenos, sendo a pele a mais significante1. A pele fornece uma superfície que é difícil de penetrar e gera uma excelente defesa contra infecções. Isto explica porque há um aumento na incidência de infecções locais e sistêmicas quando esta barreira é comprometida, por exemplo em pacientes com doenças de pele ou queimaduras.
Patógenos podem entrar no corpo através de outras áreas anatômicas. Os tratos respiratórios, gastrointestinais e genito urinários são locais em que a membrana mucosa especializada possui um importante papel na prevenção de infecções. É importante salientar, entretanto, que não há somente mecanismos físicos e químicos envolvidos.. Há fatores imunológicos essenciais que defendem contra infecções já neste nível. A Imunoglobulina A (IgA) é um anticorpo secretado pelas células da mucosa o qual se liga aos patógenos e suas toxinas para neutralizá-las e desativá-las antes que elas possam nos invadir.
CÉLULAS E PROTEÍNAS
As células e proteínas do sistema imune inato garantem a linha de defesa presente desde o nascimento. Diferente do do sistema imune adquirido, sua resposta não é altera por invasões e não possuo “memória”. Se a barreira anatômica for quebrada, patógenos poderão invadir os tecidos abaixo. Em nível tissular, o sistema imune inato garante defesas não específicas que previnem o crescimento microbiano e consequente infecção. O sistema responde a invasão com resposta celular e de proteínas do sistema complemente. O último sera discutido ao final deste tutorial.
A resposta celular1,2,3é provocada por leucócitos, os quais podem ser granulócitos (neutrófilos, eosinófilos e basófilos) ou agranulócitos (monócitos e linfócitos). O sistema inato deseja destruir os patógenos invasores rapidamente a fim de prevenir a disseminação da infecção. Uma importante habilidade destes é a fagocitose (figura 1). Este descreve o processo pelo qual a célula engloba material estranho e mata micróbios. Macrófagos e neutrófilos são as primeiras células fagocitárias do sistema imune inato, mas outras células também participam.
Macrófagos são a primeira células fagocitária e são uma das duas linhas celular que surgem da maturação de monócitos. Monócitos também se diferenciam em células dendriticas. Estas possuem atividade limitada atividade fagocitária, mas um papel essencial em ligas a resposta inata a adquirida ao apresentar o antígeno. Esta importância sera melhor discutida mais tarde neste tutorial.
Antes de fagocitar, o patógeno precisa ser reconhecido como estranho, de fora. Isto ocorre através de interações entre receptores especializados presentes na superfície celular de macrófagos com “padrões moleculares associados a patógenos” (pathogen associated molecular patterns, PAMPs, em inglês) que são expressados por micróbios. Um exemplo é a interação entre um receptor semelhando ao toll (toll-like receptor, TLR) expresso por macrófagos, que reconhece o lipopolissacarídeo (LPS) em bactérias gram negativas.
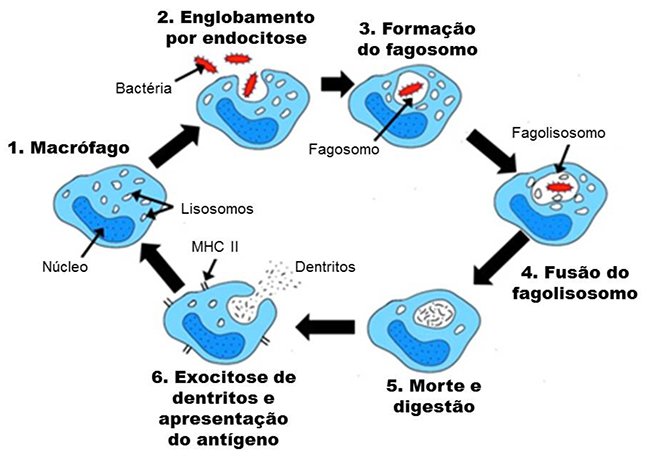
Figura 1: Interação do micróbio com o macrófago leva a fagocitose, o fagosomo se funde à lisozima do macrófago que acidifica e digere o micróbio. O macrófago apresenta partículas do micróbio em sua superfície associada ao MHC II, o que permite a ativação de outras células imunológicas.3
Uma segunda classe fagocitária são os leucócitos polimorfonucleares, mais comumente conhecidos como neutrófilos. Estes são recrutados logo emu ma infecção graças a liberação de citocinas pró-inflamatórias pelos macrófagos, como Interleucina- 1β (IL-1β), IL-6, IL-12 and IL-18. Assim aumenta-se o número de células fagocitárias no tecido que ajudam a destruir os patógenos.
Soma-se as citocinas acima citadas, macrófagos liberam fator de necrose tumoral- α (tumour necrosis factor-α, TNF-α), prostaglandinas, leucotrienos fator de ativação plaquetária. Estes mediadores induzem as mudanças vistas durante uma inflamação, tais como aumento da permeabilidade vascular, coagulação e dor. O propósito deste resposta é prevenir a disseminação da infecção por via hematológica e recrutar mais células imunes.
Nem todos os invasores são extracelulares. Vírus infectam células hospedeiras e podem evitar de serem fagocitados por macrófagos. O sistema imune inato é capaz de combater infecções intracelulares graças a outra linha células – as células natural killer (NK). Através de citocinas pró-inflamatórias, estas células são recrutadas até o local da infecção e identificam as células invadidas pela alteração ou ausência do complexo de histocompatibilidade principal (major histocompatibility complex, MHC) classe 1 (Figura 2).
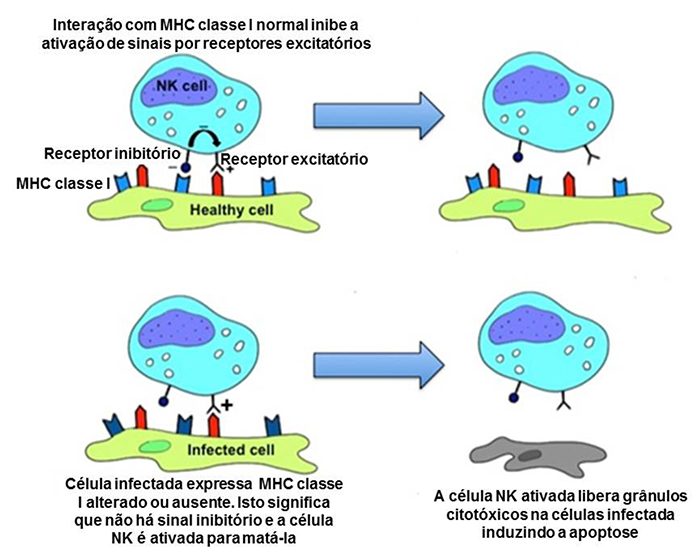
Figura 2: Células saudáveis, a imagem de cima, exibem MHC normal e não vão ativas células NK. Células infectadas, a imagem de baixo, exibem um MHC alterado ou anormal e vão ativar as células NK.1
IMUNIDADE ADQUIRIDA
A imunidade adquirida responde a infecções que foram “aprendidas” pelo sistema imunológico com o tempo. O sistema imune inato nem sempre consegue controlar uma infecção sozinho e portanto trabalha juntamente com células e proteínas altamente especializadas da resposta imune adquirida.
Após a primeira exposição do patógeno, a resposta adquirida leva um tempo para se desenvolver. Entretanto, devido a memória deste sistema, infecções subsequentes com o mesmo patógeno levam a uma resposta mais rápida. Nós usamos este sistema em nosso favor ao vacinarmos as pessoas.
A imunidade adquirida é divida em um componente humoral, os linfócitos ou células B e um componente celular, os linfócitos T ou células T. Como linfócitos, estas células surgem de células tronco hematopoiéticas na medula celular e depois são subclassificadas pelo local em que terminam de se diferenciar; células T desenvolvem-se no timo, enquanto células B na medula óssea.
Células T e B expressam receptores de superfície celular altamente específicos. O genoma dos mamíferos possui informação para milhões de receptores T e B. Durante seu desenvolvimento, células são selecionadas pela habilidade destes receptores de reconhecer antígenos específicos. Importante ressaltar que qualquer célula que responda fortemente a uma proteína própria é deletada para evitar que o sistema imune ataque o próprio corpo; este processo deve estar defeituoso em doenças autoimunes. Células que respondem fortemente a peptídeos não-próprios então permanecem virgens até que sejam ativados durante uma infecção futura em tecido linfoide secundário, como baço, linfonodos ou tecido linfoide associado a mucosa (mucosa-associated lymphoid tissue, MALT).
Células T
Células T possuem um papei na defesa do corpo contra infecções tanta extracelulares (como vírus) quanto intracelulares (como bactérias). Estas células são selecionadas pela capacidade de seu receptor de célula T (T-cell receptor, TCR) de reconhecer MHC em associação com peptídeos estranhos.
O MHC está presente em todas as células dos mamíferos, mas são específicos para cada individuo. O MHC classe I é expresso por todas as células nucleadas e o MHC classe II é expresso somente em células do sistema imunológico, predominantemente células apresentadoras de antígenos. É a interação entre o receptor de células T imaturo com o MHC I ou II que dá origem a classificação em CD4+ e CD8+. Células T imaturas expressam ambos correceptores CD8 e Cd4, os quais tem afinidade, respectivamente, por MHC classe I e por MHC classe II. Durante seu desenvolvimento no timo, as células T são expostas a peptídeos antigênicos associados ao MHC de classe I e II. Se o TCR se liga fortemente ao MHC classe, o timócito mantem o receptor CD8 e perde o CD4. Isto ocorre se o TCR possuir afinidade com o MHC classe II. Seguindo o desenvolvimento no timo, células T permanecem em tecidos linfoides secundários como CD4+ ou CD8+, onde aguardam sua ativação.
Ativação de células T virgens ocorrem em um processo chamado de ‘apresentação de antígeno’. Células dendríticas e macrófagos, envolvidos com a resposta imune inata, migram para o tecido linfático, onde apresentam o antígeno em associação com o MHC, por este motivo é que estas células também são conhecidas como células apresentadoras de antígenos (antigen presenting cells, APCs). Células T CD8+ são ativadas se o peptídeo é apresentado com MHC classe I e CD4+ são ativadas se apresentado com MHC classe II. Ativação da célula T CD8+ cria uma célula T citotóxica. Como o MHC de classe I está presente em todas as células nucleadas, se uma célula é infectada, ela irá exibir um peptídeo estranho no seu MHC. Células CD8+ específicas para este antígeno matarão estas células ao liberar vesículas citoplasmáticas na células infectada. Estas vesículas contem caspases que levam a célula à morte programada por apoptose.
Células T CD4+ tem o potencial de se diferenciarem em 4 classes: T-helper 1 (Th1), T-helper 2 (Th2), T-helper 17 (Th17) ou T-regulatórias (TReg). A formação de cada subpopulação depende da citocina presente no ambiente em que a células T é apresentada ao antígeno e cada população possui um papel diferente na inflamação. A tabela abaixo retrata a diferenciação das classes de CD4+. Ë importante notar as ações inibitórias de uma classe sobre a outra, uma vez que isto permite uma defesa efetiva contra patógenos. Repostas polarizadas podem levar a reações de hipersensibilidade, as quais serão discutidas no tutorial 2.
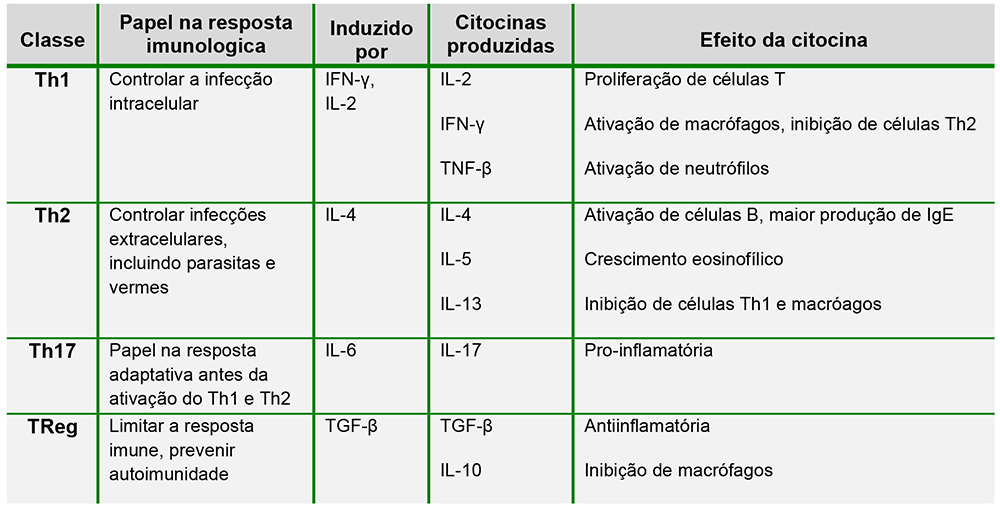
Tabela 1: Subclassificação de células CD4+1,2,4 Interferon (IFN), Fator de Crescimento Tumoral (TGF), Fator de Necrose Tumoral (TNF).
Células NKT são uma outra subclasse de células T, mais distante e independente da classificação CD4+. Estas células são capazes de reconhecer material estranho independente do MHC e libera IL-4 e IFN-γ, embora elas pareçam favorecer a resposta do tipo Th2. Não devem ser confundidas com células NK, da resposta imune inata.
Células B1,2,4
Células B estão envolvidas na resposta imune humoral, isto significa que estão envolvidas na produção de imunoglobulina (Ig) ou anticorpos e que seu papel é nos proteger contra patógenos extracelulares.
Células B exibem um receptor de células B (B-Cell receptor, BCR) altamente especifico, com afinidade para apenas 1 antígeno. Este receptor é a forma acoplada a membrana da molécula de imunoglobulina que será secretada pela célula B quando ativada. Quando o BCR interage com seu antígeno, ele internaliza o complexo receptor-antígeno, processa o antígeno e então expressa peptídeos antigênicos em associação com MHC classe II na sua superfície. Ao expressar estes peptídeos, as células B também atuam como APCs para ajudar células T que foram ativadas pelo mesmo patógeno. A célula T então libera citocinas IL-2, IL-4, IL-10 e IL-13 as quais induzem a ativação da célula B. Raramente, algumas células B podem ser ativadas sem a ajuda das células T. Isto somente ocorre na presença de antígenos raros, que podem induzir diretamente os caminhos da sinalização celular ao interagir com o BCR.
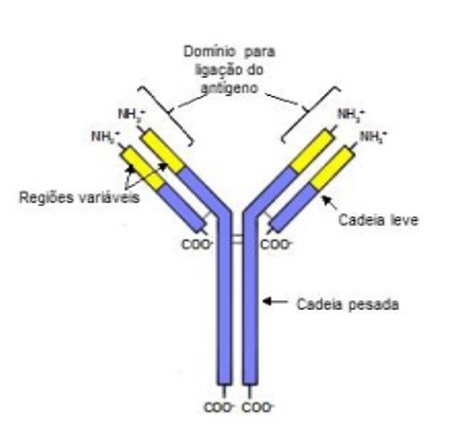
Figura 3: Estrutura da imunoglobulina5
Após a ativação, células B proliferam e se diferencial em um grande número de células idênticas, plasmáticas e de memória, em um processo conhecido como expansão clonal. Os plasmócitos são as responsáveis pela secreção de imunoglobulina, enquanto as células de memória entram em um estado de espera e se envolvem com a produção rápida e vigorosa em uma próxima infecção pelo mesmo patógeno.
Imunoglobulina 1,2,4
Imunoglobulina é um complexo proteico que consiste em duas cadeias pesadas idênticas e duas cadeias leves idênticas. No final da parte amina da molécula há um local de ligação altamente específico para o antígeno, chamada de região variável, uma vez que é nela que reside a diversidade que leva a especificidade contra diferentes antígenos. O terminal carboxi do peptídeo é conhecido como região constante é a responsável pela interação com os receptores de anticorpos presentes nos neutrófilos, macrófagos e mastócitos, além de ser capaz de induzir a cascata do complemento.
As moléculas de imunoglobulina podem ser classificas em cinco isoformas, dependendo do tipo de cadeia pesada (Tabela 2)

Tabela 2: Isoformas da Imunoglobulina4
Como mostra a tabela 2, a imunoglobulina desempenha diversos papéis na defesa do organismo, tanto intracelular quanto extracelular. O papel da IgE na ativação de mastócitos, basófilos e eosinófilos não pode ser negligenciado. IgE é produzido por células B ativadas. Embora seja a imunoglobulina menos abundante, é vital para combater infecções contra parasitas. Mastócitos, basófilos e eosinófilos são granulócitos com grânulos intracelulares contendo mediadores inflamatórios incluindo histamina, leucotrienos e TNF. Estas células expressam receptores Fc (Fc receptors, FcR) os quais são específicos para IgE. Quando o complexo IgE:FcR faz ligação cruzada com um antígeno, a célula degranula para causar a inflamação e ajudar na destruição do parasita. Este processo também contribui para alergia e hipersensibilidade, que serão melhor discutidos no tutorial 2.
Enquanto a resposta celular é essencial para uma resposta inata efetiva, um segundo mecanismo, conhecido como sistema complemento, possui um papel tão importante quanto.
Resposta do Complemento1,2,4
O fígado produz um número de proteínas do complemento envolvidas na resposta imune inata e adquirida. Aqui, a cadeia de eventos é ativada pela invasão microbiana e o processo, conhecido como cascata de ativação do complemento. Há dois caminhos principais para destruir os micróbios: opsonização e a cascata complemento.
Opsonização é a marcação dos patógenos por proteínas do complemento. Uma vez ligadas ao patógeno, as proteínas do complemento interagem com receptores específicos do complemento expresso em macrófagos e induz fagocitose. A ligação com proteína do complemento também leva a ativação da cascata do complemento. Este é um processo mais complexo para ativação proteica que diretamente induz a morte microbiana através da formação de uma ataque a membrana. Este complexo literalmente abre um buraco na parede celular e matar o micróbio.
A cascata complemento pode ser ativada por três caminhos: clássica, da lectina e alternativo. Cada caminho envolve diferentes proteínas do complemento, mas todas levam a hidrolise da proteína C3 em C3a e C3b. C3a age como uma citocina inflamatória enquanto C3b age como uma opsonina ou iniciador do caminho de ataque a membrana.
Os estágios exatos da cascata complemento são complexos e estão além da finalidade deste tutorial, entretanto, é importante lembrar de três fragmentos de proteína do complemento. C3a, C4a e C5a agem como potentes citocinas inflamatórias, causando contração do músculo liso, permeabilidade vascular e degranulação de mastócitos e basófilos. Estas proteínas são conhecidas como anafilotoxinas devido a sua habilidade de induzir sistemas anafiláticos independentemente da IgE. Anafilaxia será descrita detalhadamente no próximo tutorial.
SUMARIO
O sistema imune é um rede complexa de células que são capazes de nos proteger contra infecções. A resposta imune inata é feita pincipalmente pelas células fagocitárias e sistema complemento, enquanto o sistema imune adquirido depende de uma resposta humoral e células mais especializada originadas das células T e B. Os dois sistemas estão intimamente ligados e trabalham sinergicamente na luta contra invasores.
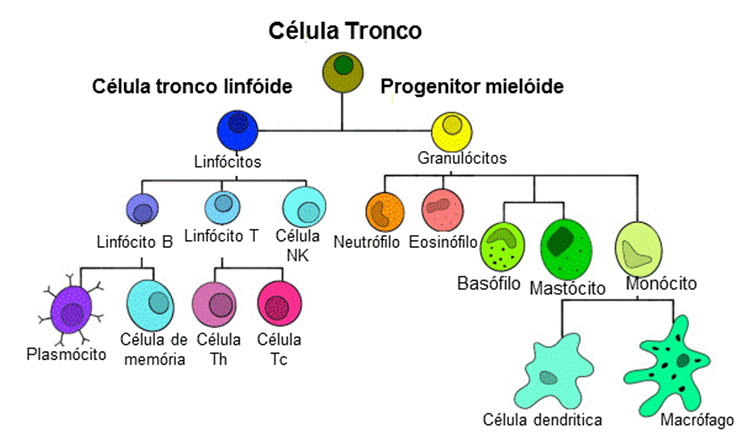
Figura 4: Células do sistema imunológico3
RESPOSTAS MCQ
- a) Verdadeira: um defeito no sistema imune mediado por células pode levar a uma inapropriada destruição das células do próprio corpo e então o desenvolvimento de um processo autoimune.
b) Verdadeira: a ativação de linfócitos B ocorre na medula óssea e dos linfócitos no timo
c) Falsa: plasmócitos são parte da resposta humoral, não do sistema imune mediado por células
d) Falsa: imunoglobulinas não estão envolvidas na resposta imune mediada por células. IgA é uma imunoglobulina de mucosa.
e) Falsa: histamia é liberada pelo mastócitos quando ativado pela IgE, o qual não pertence ao sistema imune mediado por células - a) Verdadeira: células B são assim chamadas por amadurecerem na medula óssea. As células T amadurecem no timo.
b) Verdadeira: a forma dos receptores celulares na superfície das células B é similar a das imunoglobulinas que a célula irá produzir. Isto garante que a imunoglobulina irá ligar-se ao mesmo antígeno identificado pela célula B.
c) Falsa: células NK são parte da resposta do sistema imune inato, separada da resposta humoral.
d) Falsa: Células B são ativadas principalmente por células T. Ocasionalmente, alguns antígenos são capazes de ativar as células B diretamente.
e) Falsa: cada células B produz apenas um tipo de anticorpo. - a) Falsa: imunoglobulina é produzida por plasmócitos, que são as células B.
b) Verdadeira: IgA é um tipo de imunoglobulina, encontrada nas membranas mucosas.
c) Falsa: cada imunoglobulina consiste em duas cadeias pesadas e duas cadeias leves.
d) Falsa: especificidade por antígenos depende da região variável de ambas as cadeias leves e pesadas.
e) Falsa: IgE causa degranulação dos mastócitos.
REFERENCIAS
- Murphy K, Travers P, Walport M; Immunobiology, 7th Ed. New York, Garland Science, 2008.
- Todd I, Spickett G; Lecture Notes Immunology, 6th Ed. Oxford, Wiley-Blackwell, 2010
- Todar, P. (2012). Immune Defense against Bacterial Pathogens: Innate Immunity. Available: http://textbookofbacteriology.net/innate.html Last accessed 2nd July 2015.
- Male, D; Immunology An Illustrated Outline, 3rd Ed. London, Mosby-Wolfe, 1998.
- Biochemistry Department, University of Otago. (1999). Biochemistry 111. Available: http://biocadmin.otago.ac.nz/fmi/xsl/bioc/learnbitslecture.xsl?-db=BIOCweb.fp7&-lay=Lectures&-recid=3970&-find Last accessed 2nd July 2015.
This work by WFSA is licensed under a Creative Commons Attribution-NonCommercial-NoDerivitives 4.0 International License. To view this license, visit https://creativecommons.org/licenses/by-nc-nd/4.0/